Biology Beyond Nature | ภาคภูมิ ทรัพย์สุนทร
Innovation Grand Challenges (10)
ช่วงเวลาไล่เลี่ยกับที่ Christian Anfinsen ให้ความหวังเรื่องการทำนายโครงสร้างสามมิติของโปรตีนจากลำดับอะมิโน
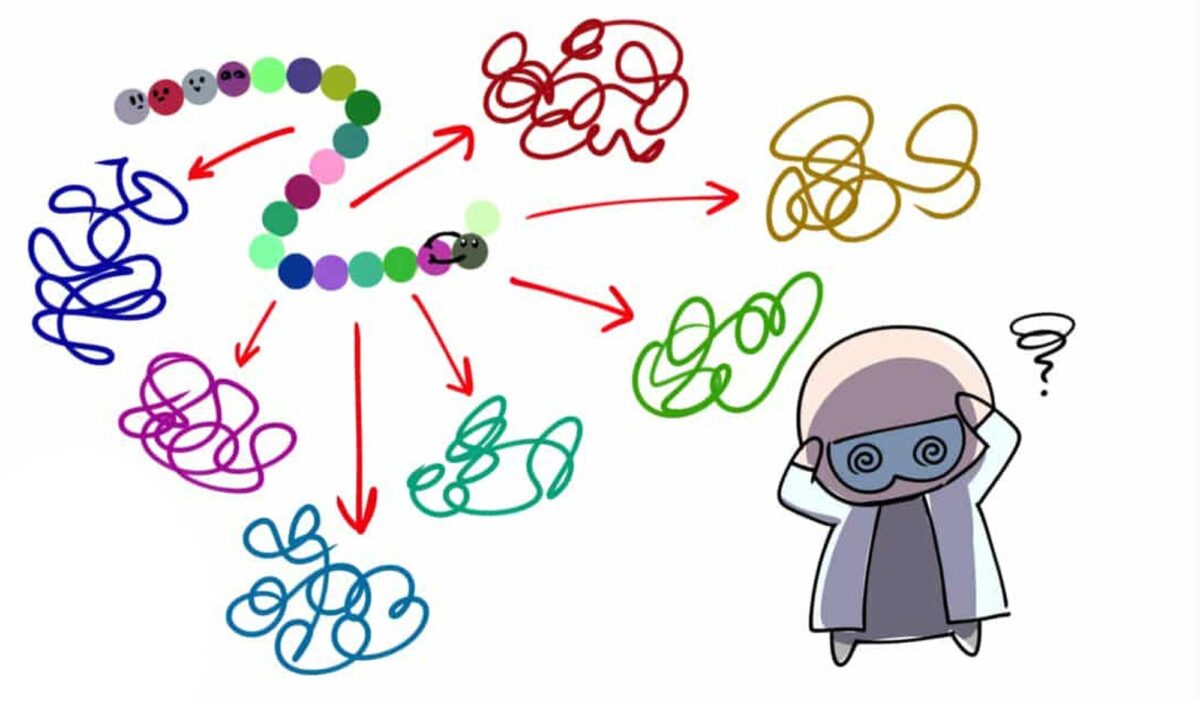
Cyrus Levinthal นักชีวโมเลกุลชาวอเมริกันเสนอปัญหา Levinthal’s Paradox แนวคิดที่ว่าแม้แต่โปรตีนสายสั้นๆ ขนาดเพียง 100 อะมิโนก็อาจม้วนพับเป็นรูปร่างในสามมิติได้ถึง 3200 แบบ
ดังนั้นเป็นไปไม่ได้เลยที่โปรตีนตามธรรมชาติจะม้วนพับแบบสุ่มไปเรื่อยๆ จนกว่าจะลงล็อกได้โครงสร้างสามมิติที่ถูกต้อง เพราะต่อให้แต่ละสามารถลองแต่ละแบบแผนได้ในเสี้ยวของเสี้ยววินาทีก็ยังต้องใช้เวลามหาศาลยาวนานกว่าอายุขัยของจักรวาลกว่าจะสำรวจความโครงสร้างที่เป็นไปได้ครบ
ส่วนในมุมของการทำนายโครงสร้างโปรตีนแปลว่าเราต่อให้เรามีแบบจำลองคอมพิวเตอร์ที่ลองทดสอบโครงสร้างสามมิติของโปรตีนแบบสุ่มไปเรื่อยๆ ในเสี้ยววินาทีต่อโครงสร้าง เราก็จะยังไม่สามารถหาโครงสร้างที่ถูกต้องได้เจอ
กลางทศวรรษ 1970-1990 หลายทีมวิจัยเริ่มพัฒนาเทคนิคทำนายโครงสร้างสามมิติของโปรตีน แบ่งได้เป็นสองแนวทางหลักๆ คือแนวทางฟิสิกส์ และแนวทางสถิติ
แนวทางฟิสิกส์อาศัยการสร้างแบบจำลองสมการของแรงต่างๆ ระหว่างแต่ละอะตอมมาทำนายว่าแต่ละส่วนของโมเลกุลโปรตีนที่เราสนใจน่าจะต้องจัดเรียงตัวอย่างไรจึงจะเสถียรที่สุด
ส่วนแนวทางสถิติอาศัยคลังข้อมูลความสัมพันธ์ระหว่างลำดับอะมิโนกับโครงสร้างสามมิติที่เรามีอยู่แล้วมาสร้างแบบจำลองทำนายโครงสร้างโดยเราอาจจะยังไม่ต้องรู้รายละเอียดทางฟิสิกส์ภายในโมเลกุล
แม้จะงานวิจัยตีพิมพ์นำเสนอเทคนิคปลีกย่อยในการทำนายโครงสร้างสามมิติของโปรตีนออกมามากมาย มีแม้กระทั่งซอฟต์แวร์สำเร็จรูปทำออกมาขาย
แต่ความก้าวหน้าที่แท้จริงของทั้งวงการยังไม่ปรากฏชัด
เหตุผลหนึ่งก็คือแต่ละกลุ่มวิจัยต่างก็เคลมว่าผลการทำนายของตัวเองดีที่สุด ภายใต้โจทย์ เงื่อนไข และวิธีการประเมินแบบที่ตัวเองเลือก เป็นเรื่องยากมากที่จะเปรียบเทียบอย่างไม่ลำเอียงว่างานชิ้นไหนดีกว่า
ที่สำคัญคือโปรตีนที่แต่ละผลงานวิจัยเหล่านี้เคลมว่า “ทำนายโครงสร้างสำเร็จ” ก็ต่างเป็นโปรตีนในฐานข้อมูลสาธารณะที่เรารู้โครงสร้างอยู่แล้ว
Cr. ณฤภรณ์ โสดา
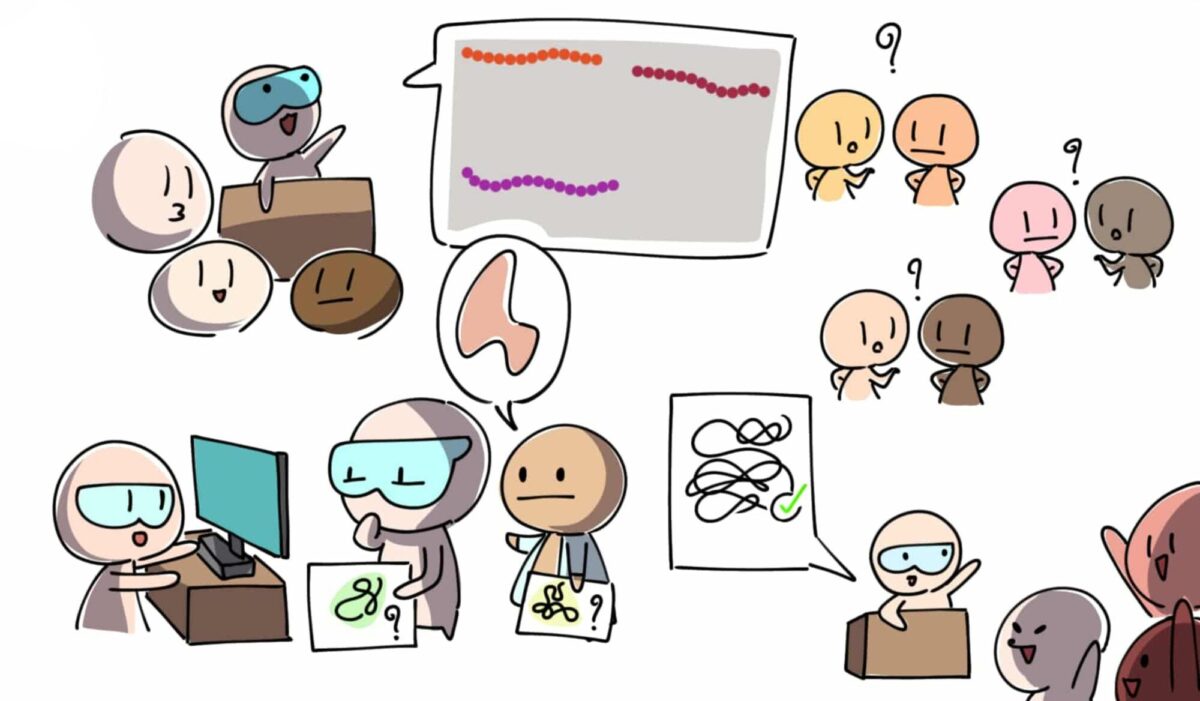
John Moult อาจารย์นักวิจัยจาก University of Maryland มองเห็นปัญหานี้และก่อตั้ง The Critical Assessment of Protein Structure Prediction (CASP) กิจกรรมการแข่งขันทำนายโครงสร้างโปรตีนที่จัดขึ้นทุกสองปีเริ่มตั้งแต่ปี 1994
กระบวนการแข่งขันเริ่มจากการเผยรายชื่อและลำดับอะมิโนของโปรตีนเป้าหมาย ทั้งทีมผู้จัดงานและผู้เข้าแข่งขันจะยังไม่รู้ข้อมูลเกี่ยวกับโครงสร้างสามมิติของโปรตีนเหล่านี้ รู้แต่เพียงว่ามีทีมวิจัยฝั่งห้องปฏิบัติการที่กำลังแก้โจทย์หาโครงสร้างสามมิติใกล้สำเร็จแล้วโดยใช้เทคนิคอย่าง X-ray Crystallography หรือ NMR
เมื่อทุกทีมที่เข้าแข่งขันส่งผลการทำนายโครงสร้างสามมิติกันมาครบ ทีมวิจัยฝั่งห้องปฏิบัติการจึงค่อยเปิด “เฉลย” โครงสร้างสามมิติที่ถูกต้อง เพื่อให้คณะกรรมการได้ประเมินว่าทีมไหนเทคนิคใดสามารถทำนายโครงสร้างสามมิติของโปรตีนได้ใกล้เคียงของจริงมากที่สุด
Moult เล่าว่าความท้าทายหนึ่งของ CASP รอบแรกคือการเชิญชวนเหล่าทีมวิจัยชั้นนำให้มาเข้าร่วมการแข่งขันซึ่งต้องลงทุนลงเวลา แถมยังต้องเอาชื่อเสียงตัวเองไปเสี่ยงหากผลงานออกมาด้อยกว่าทีมวิจัยคู่แข่งในวงการ

Cr. ณฤภรณ์ โสดา
สําหรับ CASP รอบแรก ทีมผู้จัดงานประกาศชื่อและลำดับอะมิโนให้ทีมผู้เข้าแข่งกันทราบในเดือนมีนาคม 1994 ทุกทีมมีเวลาประมาณเจ็ดเดือนในการส่งผลการทำนายก่อนผลโครงสร้างสามมิติฝั่งห้องปฏิบัติการจะเสร็จช่วงเดือนตุลาคม คณะกรรมการมีเวลาพิจารณาผลงานประมาณหนึ่งเดือน และจะเชิญผู้เข้าร่วมทั้งหมดมาประชุมดูผลร่วมกันในเดือนธันวาคม
การประชุมจัดขึ้นที่โบสถ์ไม้เก่าแก่ในศูนย์ประชุม Asilomar บนชายฝั่งแคลิฟอร์เนีย นี่คือสถานที่สำคัญทางประวัติศาสตร์ของวงการไบโอเทคที่เหล่านักวิจัยชั้นนำด้านชีวโมเลกุลเคยมาถกประเด็นอนาคตของพันธุวิศวกรรมกันเมื่อกลางทศวรรษ 1970
ที่ Asilomar เหล่าทีมนักทำนายโครงสร้างโปรตีนจะเปลี่ยนโหมดจากการแข่งขันมาเป็นความร่วมมือ เอาผลทำนายและคำตอบที่ถูกต้องมากางดูกันชัดๆ ว่าตรงไหนถูก ตรงไหนผิด และมีตรงไหนที่น่าจะปรับปรุงต่อยอดได้อีกบ้าง
เป็นบรรยากาศที่ต่างจากการประชุมวิชาการหรือตีพิมพ์งานวิจัยปกติที่ต่างทีมต่างเลือกเฉพาะผลงานที่ดีที่สุดมานำเสนอ ส่วนอะไรที่ไม่เวิร์กก็มักเก็บเงียบเอาไว้
Cr. ณฤภรณ์ โสดา
ทีมผู้จัดแบ่งโจทย์การทำนายออกเป็นสามกลุ่มตามระดับความยาก
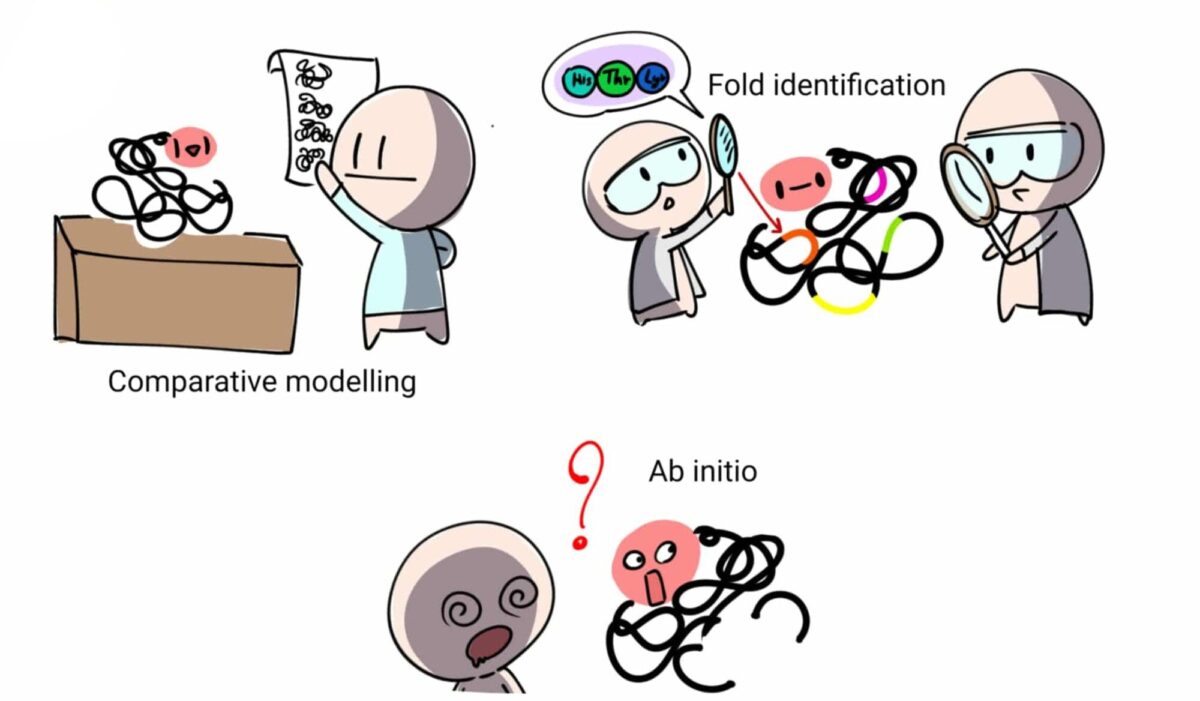
กลุ่มแรกคือกลุ่มที่เรามีโปรตีนที่เรารู้โครงสร้างสามมิติแล้วที่มีลำดับอะมิโนคล้ายคลึงกับตัวที่เราต้องการทำนาย ดังนั้นเราสามารถเอาโครงสร้างที่รู้แล้วเป็นต้นแบบเปรียบเทียบ (comparative modelling) และโฟกัสเพียงทำนายส่วนโครงสร้างปลีกย่อยที่น่าจะต่างออกไป
กลุ่มที่สองเป็นโปรตีนที่ไม่ได้คล้ายกับตัวที่เรารู้โครงสร้างอยู่เดิมแต่ว่าอาจจะมีบางส่วนบางตอนสั้นๆ ของสายอะมิโนที่อาจม้วนพับในรูปแบบใกล้เคียงกับที่เราเคยรู้จัก ดังนั้นเราสามารถจะหาส่วนม้วนพับเหล่านี้ก่อน (fold identification, Threading) มาเป็นจุดตั้งต้นในการทำนายโครงสร้าง
ส่วนโจทย์กลุ่มที่สามที่ยากที่สุดคือโปรตีนที่ดูแล้วลำดับอะมิโนไม่คล้ายกับสิ่งที่เรารู้จักโครงสร้างอยู่ก่อนเลย ดังนั้นเราแทบจะต้องเริ่มต้นจากศูนย์ (ab initio) โดยอาจจะต้องใช้แบบจำลองทางฟิสิกส์ของแรงระหว่างอะมิโนองค์ประกอบ
หรือไม่ก็ต้องใช้วิธีทางสถิติที่ซับซ้อนในการขุดหาแบบแผนของการม้วนพับในลำดับอะมิโนจากฐานข้อมูลที่อาจจะไม่ได้เหมือนกับเป้าหมายของเราอย่างตรงไปตรงมา
Cr. ณฤภรณ์ โสดา
CASP รอบแรกมีทีมเข้าแข่งขัน 35 ทีม โปรตีนเป้าหมายการทำนาย 33 ชนิด ผลลัพธ์ที่ได้คือไม่มีทีมไหนทำนายโครงสร้างโปรตีนออกมาได้ใกล้เคียงความจริงเลย
โดยเฉพาะโจทย์กลุ่มสามที่เราไม่มีโครงสร้างโปรตีนที่คล้ายคลึงเป้าหมายเป็นคู่เทียบ แม้แต่การทำนายโครงสร้างย่อยในโปรตีน เช่น การม้วนตัวเป็นเกลียว (helix) หรือจัดเรียงเป็นแถบแบน (sheet) ก็ไม่ได้ดีไปกว่าที่เคยมีการรายงานมาเมื่อสิบกว่าปีก่อน
อย่างไรก็ตามในโจทย์แบบที่สองมีตัวอย่างการทำนายที่พอใช้ได้บ้างแม้จะเป็นส่วนน้อย นอกจากนี้ยังเริ่มเห็นตัวอย่างการประยุกต์ใช้โครงข่ายประสาทเทียม (Artificial Neural Network) ในการทำนายโครงสร้าง
CASP2 จัดขึ้นในปี 1996 จำนวนทีมเข้าแข่งขันเพิ่มขึ้นมาเป็น 76 ทีม โปรตีนเป้าหมายการทำนาย 42 ชนิด รอบนี้เห็นความคืบหน้าในการแก้โจทย์กลุ่มแรกและกลุ่มสองบ้าง แต่โจทย์กลุ่มสามผลการทำนายยังคงเละเทะไม่ต่างจากรอบก่อนหน้าเท่าไร
CASP3 ในปี 1998 ทีมเข้าแข่งขันเพิ่มขึ้นมาอีกเป็น 98 ทีม โปรตีนเป้าหมายการทำนาย 43 ชนิด ข่าวใหญ่สำหรับรอบการแข่งขันนี้คือหลายกลุ่มวิจัยเริ่มทำนายโจทย์กลุ่มสามได้ใกล้เคียงความจริงขึ้นบ้างแล้ว อย่างน้อยก็ในระดับเค้าโครงโดยรวมของโปรตีนขนาดเล็ก (60-80 อะมิโน) ที่ความละเอียดของตำแหน่งอะตอม 6 อังสตรอม ความละเอียดระดับนี้ยังไม่พอจะเอาไปใช้ประโยชน์ได้ทางปฏิบัติแต่ก็ถือเป็นความก้าวหน้าสำคัญจาก CASP สองรอบก่อน
CASP3 ยังเป็นเวทีเปิดตัวอย่างเป็นทางการของ Rosetta เทคนิคทำนายโครงสร้างโปรตีนของทีม David Baker อาจารย์หน้าใหม่วัยสามสิบหกจาก University of Washington เรื่องราวของ Rosetta และ Baker จากกลายมาเป็นผู้เล่นแนวหน้าของ CASP วงการทำนายโครงสร้างและออกแบบโปรตีนอีกหลายปีต่อจากนี้
ติดตามต่อตอนหน้าครับ