ชีวิตเกิดขึ้นได้อย่างไร (บนโลกที่ไร้ชีวิต) (6) : จากห้องแล็บเคมีสู่วิศวกรผู้วิวัฒน์ไรโบไซม์

ทะลุกรอบ | ป๋วย อุ่นใจ
เลสลี ออร์เจล (Leslie Orgel) คือหนึ่งในนักเคมีระดับตำนาน เขาคือหนึ่งในเพื่อนร่วมงานที่สนิทที่สุดของฟรานซิส คริก (Francis Crick) นักวิทยาศาสตร์ชื่อดังผู้ค้นพบโครงสร้างเกลียวคู่ของดีเอ็นเอร่วมกับเจมส์ วัตสัน (James Watson) ในปี 1953
ฟรานซิสเป็นนักฟิสิกส์ ส่วนเลสลีเป็นนักเคมี
ย้อนกลับไปในช่วงปีทศวรรษ 1960 ในยุคที่นักชีววิทยากำลังมึนตึ้บกับคำถามที่ว่าโมเลกุลแรกแห่งชีวิตน่าจะเป็นโมเลกุลอะไรกันแน่ ระหว่าง ดีเอ็นเอ ที่เก็บข้อมูลกับ โปรตีนที่ทำงานเร่งปฏิกิริยาเคมี
ในตอนนั้น เรื่องนี้เป็นประเด็นใหญ่เพราะดีเอ็นเอจะก๊อบปี้ตัวเองได้ก็ต้องมีโปรตีนช่วย แต่โปรตีนจะถูกสร้างขึ้นมาได้ ก็ต้องมีพิมพ์เขียวจากดีเอ็นเอ
คำถามนี้เลยเป็นคำถามประเภท “ไก่กับไข่ อะไรจะเกิดก่อนกัน?”

แล้วถ้าคำตอบดันพลิกล็อกไม่ใช่ทั้งสองล่ะ? บางที โมเลกุลแรกแห่งชีวิต อาจจะไม่ใช่ดีเอ็นเอ หรือ โปรตีน แต่อาจจะเป็น “อาร์เอ็นเอ” เลสลี ฟรานซิส และ คาร์ล วูเซ (Carl Woese) นักชีววิทยาผู้ได้ชื่อว่าเป็นบิดาแห่งอาร์เคียได้ร่วมกันตั้งคำถาม
ไอเดียแหกคอกนี้น่าสนใจเพราะอาร์เอ็นเอทำได้ทั้งเก็บข้อมูล และสามารถเร่งปฏิกิริยาเคมี ติดตรงที่อาร์เอ็นเอนั้นมักจะไม่เสถียร สลายตัวได้ง่าย แถมจนถึงตอนนี้ก็ยังไม่มีใครเคยเจออาร์เอ็นเอที่สามารถ “ก๊อบปี้ตัวเองได้” (Self-replicating RNA) ในธรรมชาติ
ในตอนนั้น เลสลี ที่เป็นนักเคมีอินทรีย์ เชื่อว่าปฏิกิริยาเคมีที่เกิดขึ้นระหว่างสารเคมีต่างๆ ในน้ำโคลนก้นสมุทรที่เรียกว่า ซุปก่อนชีวกาล หรือ prebiotic soup นั้นน่าจะทำให้เกิด “อาร์เอ็นเอ” ขึ้นมาโดยบังเอิญ
และเพื่อพิสูจน์ เขาและทีมวิจัยของเขาที่สถาบันวิจัยซอล์ค (Salk Institute) ได้ทุ่มเททำการทดลองซ้ำแล้วซ้ำเล่าเพื่อจะพิสูจน์ว่า “เคมีนั้นอาจสร้างชีวิตขึ้นมาได้”
แต่แม้ว่าเขาจะพยายามทุ่มเททดลองสังเคราะห์อาร์เอ็นเอสักเพียงไร การทดลองของพวกเขาก็ยังไม่สามารถคอนเฟิร์มอะไรได้ชัดเจน
จวบจนถึงช่วงทศวรรษที่ 1980 เขาได้พบกับ เจอรัลด์ จอยซ์ (Gerald Joyce) นักศึกษาแพทย์ไฟแรงที่สนใจไขปริศนา “กำเนิดแรกแห่งชีวิต” เช่นเดียวกันกับเลสลี
เจอรัลด์ฉลาดหลักแหลม และมีแววเป็นนักวิทยาศาสตร์ หลังจากที่ได้คุยกัน เลสลีก็ตัดสินใจชวนเจอรัลด์เข้ามาร่วมทีมในตำแหน่งนักศึกษาปริญญาเอกเพื่อทำโปรเจ็กต์เพื่อพิสูจน์ว่าอาร์เอ็นเอนั้นน่าจะเกิดเองได้ด้วยวิธีการทางเคมีสังเคราะห์
และไม่ช้าไม่นาน เจอรัลด์ก็ทำให้เลสลีที่เป็นนักเคมีสายบริสุทธิ์ต้องแปลกใจ … และต้องเริ่มคิดใหม่ ในการสังเคราะห์สายอาร์เอ็นเอด้วยวิธีทางเคมี
เจอรัลด์เจอ “ตอ” ใหญ่เบิ้ม ในธรรมชาติ บางที เคมีเฉยๆ ก็อาจจะเอาไม่อยู่
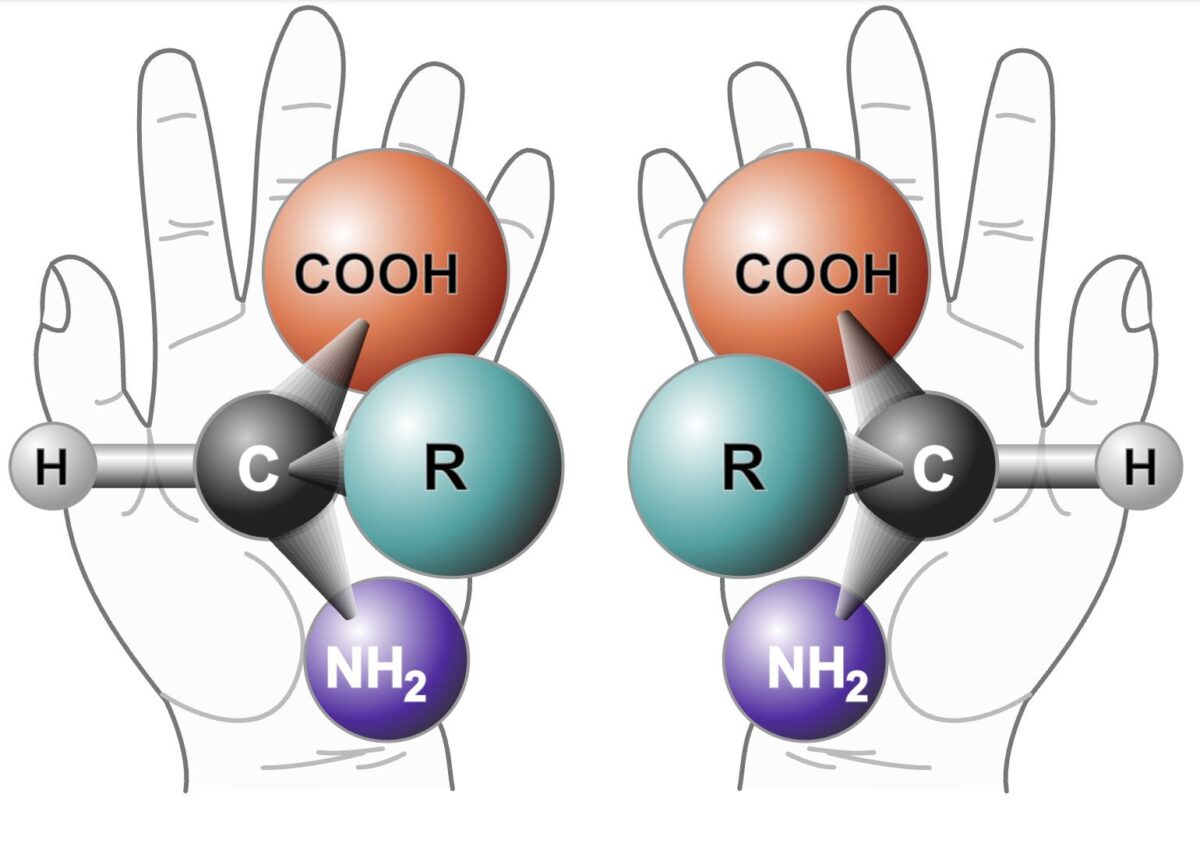
ในวงการเคมี เป็นที่รู้กันดีว่า โมเลกุลหลายชนิดรวมถึง นิวคลีโอไทด์ ที่เป็นองค์ประกอบของอาร์เอ็นเอและดีเอ็นเอ จะมีคุณสมบัติที่เรียกว่า “ไครัล (Chiral)” หรือการมีสมมาตรแบบ “ภาพสะท้อนในกระจก”
เปรียบก็เหมือนมือซ้ายกับมือขวา แม้จะดูเหมือนกันเป๊ะ แต่เอามาแทนกันไม่ได้ คุณไม่มีทางเอาถุงมือซ้ายมาใส่สวมมือขวาให้พอดีได้ฉันใด โมเลกุลในโลกเคมีก็เป็นฉันนั้น โดยนักเคมีจะเรียกภาพสะท้อนในกระจกแบบนี้ว่า D (ขวา) และ L (ซ้าย)
เจอรัลด์พบว่านี่คือจุดเริ่มต้นของหายนะของการสังเคราะห์อาร์เอ็นเอในห้องแล็บ…และในธรรมชาติ

ประเด็นก็คือในสิ่งมีชีวิตทุกชนิดบนโลก สายอาร์เอ็นเอจะถูกสร้างขึ้นจากนิวคลีโอไทด์แบบ D (ขวา) เท่านั้น แต่ในซุปขี้โคลนธรรมชาติ หรือแม้แต่ในการทดลองเคมีแบบสุ่มของเจอรัลด์ สารเคมีที่พบนั้นดันมีทั้งแบบซ้าย (L) และขวา (D) ผสมกันแบบละครึ่ง ซึ่งทำให้มีโอกาสราวๆ 50-50 ที่สายอาร์เอ็นเอสร้างใหม่จะไปเอานิวคลีโอไทด์ที่กลับข้างมาใช้
และเมื่อโมเลกุล “L” (ซ้าย) เข้าไปต่อในสายอาร์เอ็นเอ โครงสร้างที่ผิดด้านของมันจะเข้าไปขวาง ทำให้การต่อสายอาร์เอ็นเอหยุดชะงัก เหมือนการต่อเลโก้ หากมีตัวต่อที่สลับด้านโผล่มาเสียบ การต่อเพิ่มบางทีก็ไปต่อไม่ได้ (กระบวนการนี้ในทางเคมีเรียกว่าเกิดการยับยั้งแบบไขว้ของโมเลกุลภาพสะท้อนในกระจก หรือ Enantiomeric cross-inhibition)
เหตุการณ์นี้ทำให้นักเคมีสายบริสุทธิ์อย่างเลสลีถึงกับต้องเริ่มคิดใหม่… ว่าบางทีแค่ “เคมี” และ “ความบังเอิญ” อย่างเดียว อาจจะไม่เพียงพอที่จะทำให้ชีวิตถือกำเนิดขึ้นมาได้จริงๆ แม้ว่าเลสลีจะยังปักใจเชื่อว่า ลำพังเพียงปฏิกิริยาเคมีก็น่าจะเพียงพอต่อการถือกำเนิดของอาร์เอ็นเอโมเลกุลแรกและยังหาทางแก้คำตอบในเรื่องประเด็นไครัลในทางเคมี
แต่เจอรัลด์นั้นกลับเริ่มมองต่างออกไป ในมุมมองของเจอรัลด์ คำตอบของปัญหาไครัล ก็คือ ตัวเร่งปฏิกิริยา ซึ่งอาจจะเป็น “ไรโบไซม์”
เจอรัลด์ และเลสลี ตีพิมพ์ผลงานวิจัยของพวกเขาออกมาในวารสาร Nature ในปี 1984
สำหรับเจอรัลด์ การค้นพบของไรโบไซม์ของโทมัส เช็ค (Thomas Cech) ที่มหาวิทยาลัยโคโลราโด (University of Colorado) และการคัดแยกอาร์เอ็นเอที่ก๊อบปี้ตัวเองได้ (แม้จะแค่บางส่วน) ของเดวิด บาร์เทล (David Bartel) และแจ็ก โซสแต็ก (Jack Szostak) ที่โรงพยาบาลแมสซาชูเซตส์ เจเนรัล (Massachusetts General Hospital) คือ ช่วงเวลายูเรก้า…
เดวิดคือคนที่พิสูจน์ว่า อาร์เอ็นเอ ทำงานได้ดีและมีประสิทธิภาพในการเร่งปฏิกิริยาได้ไม่แพ้โปรตีน เดวิดค้นพบไรโบไซม์มากมายหลายกลุ่ม และแบ่งออกเป็นคลาสต่างๆ ที่แตกต่างกันจากการจำลองวิวัฒนาการในหลอดทดลอง (in vitro selection) เขาเชื่อว่าไรโบไซม์น่าจะมีโครงสร้างที่หลากหลายมากกว่าที่เราจินตนาการ ถ้าเราตั้งใจเฟ้นหาจริงๆ …
ด้วยแรงบันดาลใจและรากฐานมาจากงานของเดวิด เจอรัลด์ และเจมส์ โรเจอร์ (James Rogers) ได้นำเอนไซม์ของเดวิดมาท้าทายขีดจำกัดทางวิวัฒนาการ พวกเขาทดลองบีบให้ระบบสร้างไรโบไซม์โดยใช้วัตถุดิบเพียงแค่ 3 นิวคลีโอไทด์ (A, G, U) โดยบังคับตัดตัวอักษร C (Cytidine) ออกไปอย่างสิ้นเชิง (เพราะ C ไม่ค่อยเสถียร อาจจะสลายไปเป็นเบส U ได้)
ผลลัพธ์ที่ได้คือไรโบไซม์ตัวใหม่ที่ชื่อว่า R3 แม้ความสามารถในการเร่งปฏิกิริยาของมันจะช้าลงอย่างเห็นได้ชัดเมื่อเทียบกับเวอร์ชั่นที่มีเบสครบสี่ตัว
แต่งานชิ้นนี้ก็เป็นหลักฐานชั้นดีที่ยืนยันว่า ในช่วงต้นกำเนิดของชีวิต แม้องค์ประกอบทางเคมีจะขาดๆ เกินๆ ไรโบไซม์ก็ยังสามารถประกอบร่างและเกิดขึ้นได้
ต่อมา เพื่อดูว่าเอนไซม์จะพัฒนาศักยภาพไปได้ไกลแค่ไหนหากข้อจำกัดถูกปลดล็อก
เจมส์จึงทดลองนำ R3 มาวิวัฒนาการต่อ โดยอนุญาตให้มีเบส C กลับเข้ามาในระบบได้อีกครั้ง
ผลที่ได้คือเอนไซม์ตัวใหม่ที่ชื่อว่า R3C สิ่งที่น่าทึ่งคือ R3C ไม่ได้ใช้เบส C พร่ำเพรื่อ แต่มันเลือกดึง C เข้าไปแทรกในโครงสร้างเพียงไม่กี่ตำแหน่ง ซึ่งเพียงพอที่จะพลิกโฉมโครงสร้างสามมิติของมันให้กะทัดรัดขึ้น และเพิ่มประสิทธิภาพในการเร่งปฏิกิริยาให้รวดเร็วกว่าเดิมถึงกว่า 20 เท่า
การศึกษาของเจมส์ทำให้เจอรัลด์และทีมเริ่มตื่นเต้น เพราะผลการวิจัยนี้บอกอะไรเราเยอะเกี่ยวกับการดีไซน์ไรโบไซม์
และเพื่อออกแบบไรโบไซม์ใหม่ให้สามารถควบคุมได้ (programmable) เจอรัลด์ให้ลูกศิษย์ของเขา นาตาชา พอล (Natasha Paul) เริ่ม “ชำแหละไรโบไซม์” และมองมันผ่านสายตาของ “วิศวกร” ว่า ชิ้นส่วนไหนในโมเลกุลนี้ที่เป็น “หัวใจหลัก” ในการเร่งปฏิกิริยา และชิ้นส่วนไหนที่เป็นเพียง “ส่วนเกิน” ที่ไม่ได้มีความสำคัญอะไร
และที่สำคัญ ไรโบไซม์ที่เดวิดค้นพบในการคัดแยกของเขา เป็นไรโบไซม์แบบ cis-acting นั่นคือมันจะทำปฏิกิริยากับตัวเอง เช่น ต่อหรือเชื่อมสายของตัวเอง ซึ่งสำหรับเจมส์ ไม่ค่อยเป็นเหตุเป็นผลเท่าไร สำหรับอาร์เอ็นเอที่จะเป็นโมเลกุลแรกแห่งชีวิต ก็ถ้าไรโบไซม์ทำหน้าที่เร่งปฏิกิริยาได้แค่ต่อสายตัวเองแล้วจบ แล้วจะไปเป็นต้นแบบก๊อบปี้อาร์เอ็นเอลูกหลานได้ยังไง…
นาตาชาจึงจัดการ “ผ่าตัด” วงจรการทำงานเสียใหม่ โดยออกแบบให้มันทำงานแบบ trans-acting อย่างสมบูรณ์แบบ ไรโบไซม์ของเธอจะไม่ยอมให้ชิ้นส่วนไหนมาเชื่อมติดกับร่างกายตัวเองอีกต่อไป แต่มันจะทำหน้าที่เป็นแม่พิมพ์ (Template) คอยดึงเอาชิ้นส่วน RNA สั้นๆ 2 ชิ้นที่ลอยอยู่ในน้ำเข้ามาประกบ แล้วเร่งปฏิกิริยาเชื่อมชิ้นส่วนทั้งสองให้ติดกัน
ความอัจฉริยะของระบบของนาตาชาก็คือ ชิ้นส่วน 2 ชิ้นนั้น ก็คือลำดับเบส “ครึ่งหน้าและครึ่งหลัง” ของตัว R3C นั่นเอง!
ดังนั้นเมื่อเชื่อมกันเสร็จ มันจึงกลายเป็นร่างโคลนของแม่แบบอย่างสมบูรณ์”!
และแล้ว นาตาชาก็ทำได้สำเร็จ! งานของเธอปลดล็อกให้ R3C สามารถดึงชิ้นส่วนมาประกอบกัน จนกลายเป็นร่างโคลนของตัวเอง (Self-replication) ได้ในหลอดทดลอง ซึ่งเป็นการช่วยพิสูจน์ให้เห็นเป็นประจักษ์ว่าอาร์เอ็นเอสามารถถูกออกแบบให้ต่อสายพันธุกรรมเพื่อจำลองตัวเองได้จริง นี่คือการปูทางครั้งประวัติศาสตร์สู่การสร้างระบบชีวิตเทียมอย่างที่ไม่เคยมีใครทำได้มาก่อน
แต่ความสำเร็จนี้ก็มาพร้อมกับอุปสรรคชิ้นโต แม้นาตาชาจะหวังให้ระบบทำงานเป็นวัฏจักร (ดึงมาเชื่อม ปล่อยตัวลูก แล้วไปจับคู่ใหม่) แต่ในความเป็นจริง อาร์เอ็นเอรุ่นลูกหลานที่เพิ่งเกิดใหม่กลับทำงานได้ไม่ดีนัก
สาเหตุหลักคือ ตัวลูกมักจะพับตัวแบบผิดๆ ถูกๆ จนบางครั้งก็พับผิดรูป (Kinetic traps) จนสูญเสียโครงสร้างสามมิติที่เหมาะสมในการเร่งปฏิกิริยา ทำให้ไรโบไซม์ทำงานได้แย่ลง
และที่เลวร้ายกว่านั้น เนื่องจากตัวลูกถูกสร้างขึ้นบนตัวแม่ ลำดับเบสของพวกมันจึงเป็นคู่สมกันเป๊ะๆ ทำให้เมื่อสร้างเสร็จ พวกมันจึงยึดติดกันแน่น ติดหนึบ ไม่ยอมหลุดออกจากสายอาร์เอ็นเอแม่แบบเพื่อไปทำหน้าที่ของตัวเองต่อ ปฏิกิริยาจึงหยุดชะงักและไม่เกิดการเพิ่มจำนวนแบบทวีคูณ (Exponential growth) ตามที่ตั้งใจไว้
อย่างไรก็ตาม นาตาชาก็ยังตีพิมพ์งานวิจัยของเธอในวารสาร Proceedings of the National Academy of Sciences (PNAS) เรื่อง A self-replicating ligase ribozyme ในปี 2002
แค่นี้ก็น่าตื่นเต้นมากแล้ว แม้ว่าผลที่ได้จะยังไม่เพอร์เฟ็กต์ สำหรับเจอรัลด์ ปัญหาที่นาตาชาพบทั้งเรื่อง “การพับผิด” และ “การติดหนึบ” เป็นเหมือนหอกข้างแคร่ที่คอยทิ่มแทงอยู่ในใจ
และเพื่อจัดการกับปัญหานี้ เจอรัลด์ได้มอบหมายให้ เทรซี่ ลินคอล์น (Tracey Lincoln) นักวิจัยอีกคนในทีม มารื้อระบบเสียใหม่ และเริ่มออกแบบในมุมที่แตกต่างออกไป
เทรซี่ตัดสินใจดัดแปลงไรโบไซม์ให้เป็นสองเวอร์ชั่น และให้พวกมันทำงานสลับกันประกอบร่างให้อีกฝ่าย (สมมุติว่าเป็น E1 และ E2 โดยที่ไรโบไซม์ E1 ก๊อบปี้สายลูกของ E2 ในขณะที่ไรโบไซม์ E2 ก็ก๊อบปี้สายลูกของ E1) เรียกว่าระบบ จำลองแบบไขว้ (Cross-replication)
การทำเช่นนี้ทำให้คู่เบสตรงรอยต่อสลับตำแหน่งกัน ช่วยทำลายความสมมาตรที่เคยทำให้แม่และลูกจับกันแน่นเกินไปจนผลผลิตติดหนึบอยู่กับแม่แบบ ช่วยให้ผลผลิตหลุดออกจากไรโบไซม์แม่แบบได้ง่ายขึ้น
และนั่นจะทำให้ไรโบไซม์พวกนี้สามารถจำลองแบบตัวเองแบบเพิ่มทวีคูณ ได้อย่างที่ควรจะเป็นจริงๆ ไม่ต้องเจอปัญหาสายลูกติดหนึบกับสายแม่อีกต่อไป
และนี่คือจุดเริ่มต้นที่ทำให้โมเลกุลเหล่านี้สามารถเกิด วิวัฒนาการแบบดาร์วิน (Darwinian evolution) แข่งขันแย่งชิงกันได้จริงๆ ในหลอดทดลอง…
จำลองภาพปฐมบทแห่งชีวิตเหมือนหลายพันล้านปีก่อน ก่อนที่จะวิวัฒน์ต่อมาจนเป็นโลกที่มีความหลากหลายมากมายมหาศาลแบบที่เราเห็นได้ในปัจจุบัน!!!